










| Esperó de Formentera |
 |
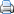 |
 |
| dimarts, 27 de novembre de 2007 11:12 | |||

Delphinium pentagynum Lam. subsp. formenteranum N.Torres, L. Sàez, Rosselló & C Blanché, Bot. J. Linn. Soc. 133: 371-377 (2000).
L'esperó de Formentera és un tàxon de descripció recent, descobert a l'illa de Formentera, d'on és endèmic. És conegut d'una única localitat, on té diversos nuclis poblacionals petits i un nombre d'efectius molt baix. Es troba amenaçat per la pressió de construcció i urbanització i pels freqüents incendis de la vegetació circumdant. Descripció i identitat taxonòmica Descripció Planta vivaç, amb soca tuberosa i fibrosa. Tiges de 38-55 cm, poc ramificades. Fulles basals palmatipartides. Raïm florífer lax, amb 5-7 flors. Flors d’un blau-violat, de 17-19 mm, amb esperó de 8-10 mm. 5 fol·licles, de 7-9 mm, pubescents. Granes d’un bru fosc a negrós, subpiramidals, cobertes d’esquames estretes.Tàxon diploide amb 2n=16 cromosomes (Torres et al., 2000; Castro & Rosselló, en premsa) Afinitats taxonòmiques i origen Es diferencia de la subspècie típica (subsp. pentagynum) per les dimensions més reduïdes de tota la planta i, en especial, per la inflorescencia pauciflora i més laxa i per les flors de mida clarament més petita, de manera que hom pot assimilar les poblacions de la subsp. formenteranum com un esquizoendemisme que comparteix el mateix nombre cromosòmic (2n=16), vicariant illenca adaptada a condicions de sequedad i d’insularitat, derivada de poblacions ibèriques o africanes (sense poder-ne precisar l’origen amb les dades disponibles en l’actualitat, cf. Torres et al., 2000) de les quals s’hauria diferenciat gràcies a l’aïllament geogràfic.
Mapa de distribució de D. pentagynum subsp. pentagynum (ratllat) i subsp. formenteranum (triangle), segons Torres et al. (2000)
Experimentalment (Bosch, 1999) s’han obtingut híbrids de caràcters intermedis, a partir d’encreuaments amb espècies properes de la ser. Pentagyna B. Pawl., que demostren l’existència de barreres genètiques febles i de divergència genètica també petita i que suggereixen que el model de diversitat del grup es basa mecanismes d’aïllament de tipus geogràfic, fenològic o ecològic (com ara la insularitat). En aquest context, es troba suport a la hipotesi que la subsp. formenteranum s’hauria originat per aïllament del conjunt de les poblacions ibèrico-nordafricanes de D. pentagynum s.l.)
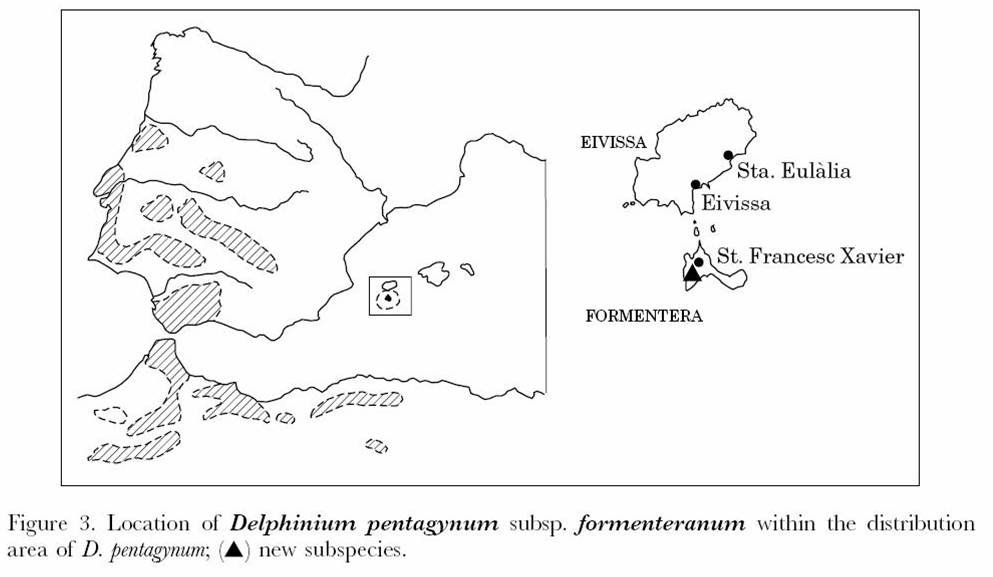
Distribució geogràfica Endemisme de les Pitiüses (Formentera). Restringit inicalment (Puget et al., 1995; Torres et al., 2000) a la capçalera del Torrent de Cala Saona, on hi ha uns pocs nuclis poblacionals. Darrerament, Castro & Rosselló (en premsa) han indicat una nova població (N. Torres & J.A. Rosselló legerunt) prop de la localitat típica, en brolla calcícola sota coberta de Pinus halepensis i, a les fitxes-recull del Projecte BioAtles (2007), ja s’indica de 8 quadrats UTM de 1 x 1 km de costat a Formentera, des de la zona d’es Cap fins a la base de la Mola, de manera que tindria una àrea superior a la que es pensava, segons dades del mapa de B. Klahr.
Quadrats UTM amb presència de l’esperó de Formentera, segons mapa de B. Klahr (in BioAtles, 2007), redibuixat sobre la imatge de software lliure http://commons.wikimedia.org/wiki/Image:Formentera_sat.png
Hàbitat
Clarianes dels fragments romanents de la màquia de Pistacia lentiscus i Juniperus phoenicea, de vegades amb Pinus halepensis, entre la brolla de Juniperus oxycedrus i Rosmarinus officinalis i en pastures seques d’aquestes clarianes, dominades per Brachypodium retusum, sobre materials calcaris. 20-50 m s.m
Individu en flor a les clarianes de la màquia amb pins del centre de l'illa, al maig de 2002 (Foto: C. Blanché)
Reproducció i estratègia vital Vegetativa: La producció de rizomes permet un creiexement clonal vegetatiu limitat que, tanmateix, assegura la pervivència de les poblacions en el temps i en l’espai. Pot representar una estratègia alternativa de manteniment per fer front a canvis ambientals excepcionals (sequeres, predació, manca temporal de pol·linitzadors) Sexual: Hermafrodita, entomòfila especialitzada i al·logama autocompatible. Sense problemes aparents i amb bones taxes de producció de granes. Pol·linització. Presenta flors proteràndriques (en què els estams maduren abans que els estigmes siguin receptius). Les flors presenten recompenses abundants (nèctar i pol·len) que ofereixen atracció als insectes pol·linitzadors, que les visiten afavorint la pol·linització encreuada. Els vectors pol·linitzadors legítims aterren a la superfície generada per l’extensió dels sèpals inferiors i, forçant la separació dels pètals laterals que actuen com a cortina, atenyen el nèctar contingut al fons de l’esperó. En aquest moviment, la regió ventral del cos de l’insecte entra en contacte amb els òrgans reproductors per recollir el pol·len (en la primera fase, masculina, els grans de pol·len s’adhereixen al cos de l’insecte) o per alliberar-lo (en la segona fase, femenina, que té lloc 1-3 dies després del marciment dels estams, on els estigmes receptius i oberts es troben en la posició exacta on recullen els grans adherits a la regió ventral). D’aquesta manera, és impossible que el pol·len es dipositi als estigmes de la mateixa flor, evitant l’autopol·linització atès que ambdues fases es troben separades en l’espai i en el temps (dicogàmia i hercogàmia); no obstant, pot existir un petit solapament, suficient per a la producció de granes en absència de pol·linitzadors (5 % d’autopol·linització espontània en experiments d’embossament experimental de flors, dades inèdites), que actuaria com a mecanisme de seguretat alternatiu.Un cop formades les granes, es dispersen per gravetat i boleoautocòria, aprofitant l’efecte de catapulta del vinclament de les tiges fructíferes (a causa del vent, del trepig dels animals, etc.), on a l’extrem, els fol·licles secs s’han obert per la sutura ventral i permeten la sortida de les llavors. Floració: V-VI – Fructificació: VI-VII
Individu en floració, maig de 2002. (Foto: C. Blanché)
Diversitat genètica
Ha estat estudiada amb tècniques de diversitat al·loenzimàtica (López-Pujol et al., 2003), que han detectat nivells moderats de diversitat (P= 40,5%; A= 1.6 i He= 0,180), equivalents als reportats per a d’altres esperons (Delphinium) de característiques biològiques similars i superiors als de la majoria d’endemismes insulars. Pot contrubuir al manteniment d’aquests valors de diversitat l’existència d’un estratègia de rizomes dorments, que mantindrien determinats genotips d’un any per l’altre, variant a l’alça els valors de Ne (nombre real efectiu disponible d’individus interfèrtils, més gran al llarg dels anys, per emergència de rizomes temporalment latents, c. 50% anual), així com el contacte amb d’altres poblacions properes. Igualment s’han trobat nivells de consanguinitat moderats, probablement a conseqüència de l’estructura genètica de les poblacions (biparental inbreeding), que s’estructuren en nuclis poblacionals que presenten riquesa al·lèlica correlacionada amb el cens d’individus reproductius del rodal. Els fragments més petits tenen una dràstica erosió genètica, la qual cosa recomana mantenir la mida poblacional suficient. No sembla haver-hi impediments al flux gènic entre rodals propers. Demografia No s’ha realitzat un seguiment demogràfic sistemàtic continuat en els anys. Les dades d’observacions de camp dels anys 1999, 2001, 2003 i 2004 indiquen l’existència de 3 nuclis poblacionals (un d’ells amb només 3 individus) on les plantes se situen en rodals o veïnatges genètics. Les plantes produeixen rosetes de fulles basals (finals d’abril); el percentatge de rosetes que arriba a florir és del 40-50% en els anys observats, mentre que la resta entren en període de repòs (en d’altres endemismes mediterranis de Delphinium, no s’ha trobat cap relació entre nivells de diversitat genètica o d’heterozigosi i paràmetres de fittness com ara les taxes de floració; els percentatges de rosetes florides podrien deure’s més aviat a la disponibilitat de recursos cf. Orellana et al., 2007). Hi ha un percentatge de rizomes dorments/latents no avaluat. Els censos més detallats que coneixem corresponen a l’any 2001, on van ser identificats 249 adults reproductors corresponents a un total de 480 rosetes foliars, cobrint una superfície de 0,75 quilòmetres quadrats (àrea d’ocupació). Desconeixem els resultats dels censos encarregats en els darrers anys pel Govern Balear.
Conservació Amenaces
Avaluació
Protecció legal
Mesures de conservació proposades (López-Pujol et al., 2004)
Situació actual (2007) Indicada com a en “moderada regressió” (Projecte BioAtles, 2007). No existeix cap pla de recuperació aprovat sobre el tàxon, ni de protecció sobre l’espai o l'hàbitat que ocupa. Afectació de la localitat clàssica per la instal·lació d'un parc de plaques d'energia solar (v. notíciaa la pàgina principal, de BioC, octubre de 2007). El Servei de Protecció d'Espècies de la Conselleria de Medi Ambient del Govern Balear ha autoritzat l'actuació amb l'adopció de mesures correctores (delimitació física de la parcel·la, prohibició de maquinària pesant i d'herbicides, seguiment per part de tècnics, etc.) i l'estudi de camp corresponent (maig de 2007) que ha comportat el reconeixement de diverses localitats addicionals a l'illa on s'ha detectat la presència de l'esperó de Formentera (cf. mapa anterior) Bibliografia Bosch, M. (1999). Biologia de la reproducció de la tribu Delphiniae a la Mediterrània Occidental. Arxius de la Secció de Ciències, 120. Institut d’Estudis Catalans, Barcelona. Lopez-Pujol, J., M. Bosch, J. Simon & C. Blanché (2003). Population genetics and conservation priorities for the critically endangered island endemic Delphinium pentagynum subsp. Formenteranum. Biodiversity and Conservation 12: 1937-1951. López Pujol, J., M. Bosch, J. Molero, A. Rovira, J. Simon & C. Blanché (2004). Delphinium pentagynum subsp. formenteranum N. Torres, L. Sàez, Rosselló & C. Blanché. In: A. Bañares et al. (eds.), Atlas y Libro Rojo de la Flora Vascular Amenazada de España, 2ª ed. . Ed. Dirección General de Conservación de la Naturaleza, Madrid: 216-217. Orellana, M.R., J. López-Pujol, C. Blanché & M. Bosch (2007). Relationships between heterozygosity and fitness in the Iberian threathened larkspur Delphinium bolosii. Flora 202: 161-168 Projecte BioAtles (2007). Delphinium pentagynum subsp. formenteranum in Projecte BioAtles, Servei de Protecció d’Espècies. Fitxa col·leccionable, 2 pag. Text de L. Sàez i C. Blanché; Mapa de B. Klahr. Edició Agost de 2007. Conselleria de Medi Ambient, Govern de les Illes Balears, Palma. Puget, G., M. Stafforini & N. Torres (1995). Notes florístiques de les Illes Balears, V. Bolletí de la Societat d’Història Natural de les Illes Balears 38 :63-74 Sáez, L. & J.A: Rosselló (2001). Llibre Vermell de la Flora Vascular de les Illes Balears. Servei de Biodiversitat. Conselleria de Medi Ambient, Govern de les Illes Balears, Palma. Torres, N., L. Sàez, J.A. Rosselló & C. Blanché (2000). A new Delphinium from Formentera (Balearic Islands). Botanical Journal of the Linnaean Society 133: 371-377 Citació / pdf / handle
|
|||
| Darrera actualització de dijous, 29 de setembre de 2011 09:36 |








{kind=link}